― はじめに
私が卒研生として研究室の扉を叩いたその年は、ヒトの性決定遺伝子SRYが同定された年でした。同時にその年は、ショウジョウバエの性分化が一連の遺伝子の選択的スプライシングカスケードにより制御されることが解明された年でもありました。たった一つの遺伝子の働きで性ががらりと変わること、選択的スプライシングという分子レベルの切り換えが個体レベルの性の切り換えと連動すること、私はそこに性差形成機構の魅力を感じました。東京農工大学農学部の卒研生当時に扱った研究材料はカイコでした。カイコの性はW染色体のFemと名付けられた遺伝子座により決まることが知られていました。ならばカイコのFemを同定してやろう。そう目標を定めました。卒研時代にカイコのZ染色体に座乗する卵サイズ決定遺伝子に関する研究を行った後、東京大学農学系研究科の大学院に入学後もカイコのZ染色体の新規遺伝子の同定や、その遺伝子量補正に関する研究を行いました。学位取得後は、比較ゲノム解析により性分化遺伝子のカイコオルソログを同定し、遺伝子組換え技術によりその機能を証明しました。さらに生化学的手法により新奇性決定遺伝子を発見し、カイコが他の生物には見られない固有の性決定機構をもつことを明らかにしました。本専攻の准教授に着任後、ついに念願のカイコのFem遺伝子の同定に成功しました。Femの正体はわずか20塩基程度のpiRNAと呼ばれる小分子RNAであり、piRNAが性決定を支配するとの発見は当時注目を集めました。カイコ固有の性決定遺伝子を次々と明らかにした私の研究成果は、性差形成機構に多様性があることを示唆した先駆的事例です。その後、過去類を見ない性差形成機構や新奇性決定遺伝子が相次いで報告され、性差形成機構は予想以上の多様性を示すことが明るみとなりました。全ての多細胞生物にとって普遍的な性を形成するメカニズムになぜ驚異的な多様性がみられるのか。それはおそらく、性差形成機構が単に性を形作るために必要なだけでなく、他にも重要な意義を持つからに違いありません。ではその意義とは何なのか?私達は今、この謎を解き明かすための研究を進めています。
鈴木雅京

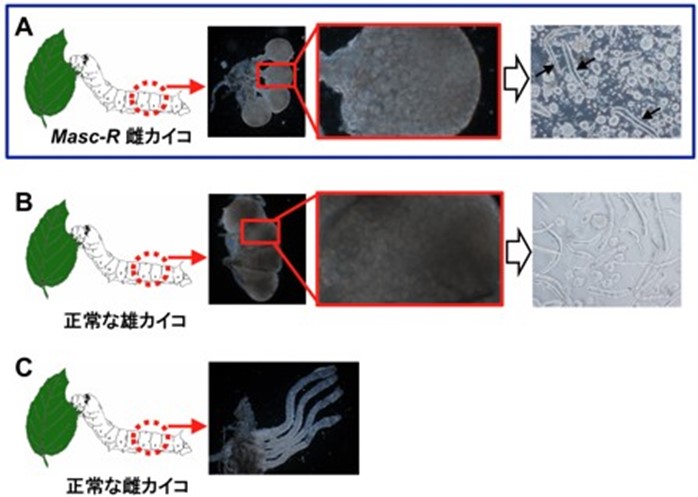
メスからオスへの性転換に成功 (Sakai et al., 2016)
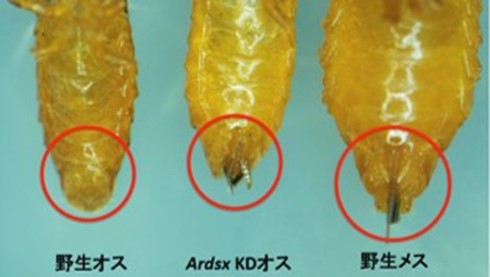
オスの生殖器がメス型に転換 (Mine et al., 2017)
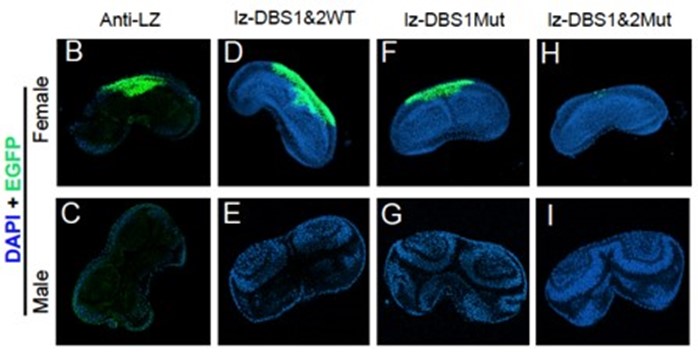
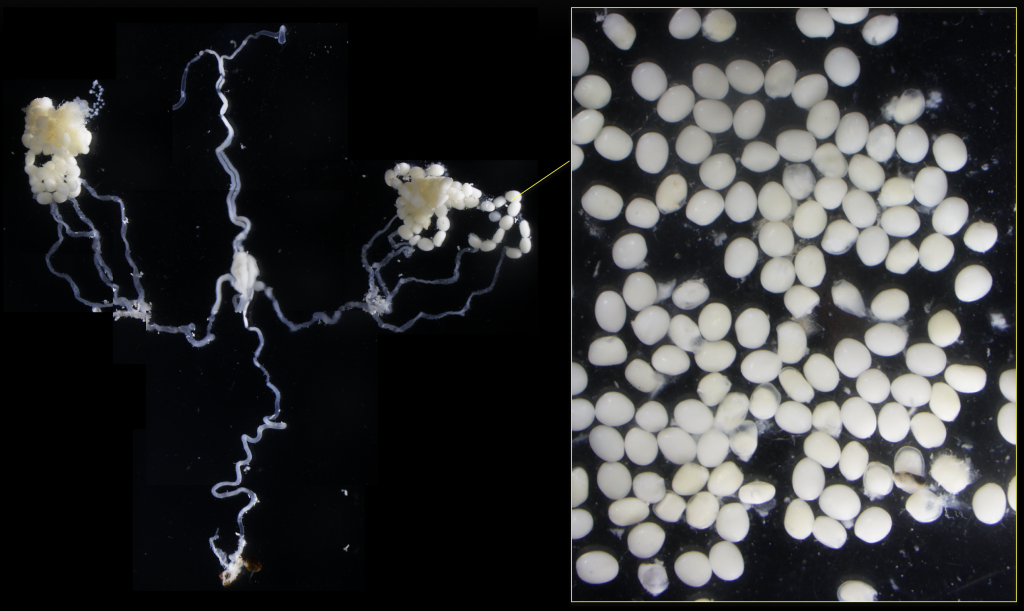
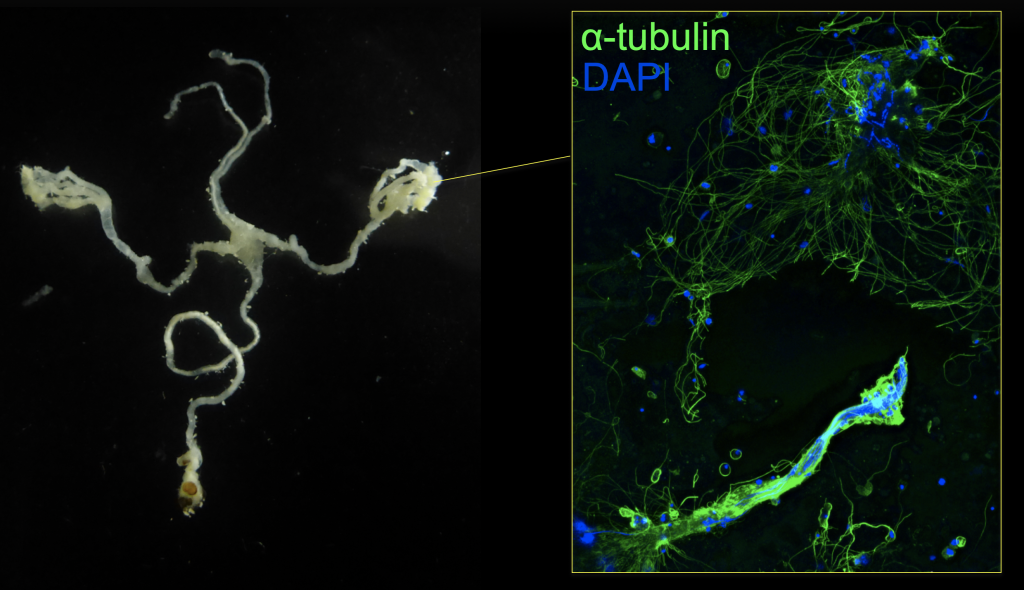
オス個体で卵、メス個体で精子が形成された
(Yuzawa et al., 2021)
― 昆虫性決定機構の多様性と昆虫特異機能に関する研究
少なからぬ人間にとって恋愛は最も魅力的なものであり、理想の相手に出会うことは人生最大の関心事といっても過言ではないでしょう。一方で特に男女関係においては、「夫婦喧嘩は犬も食わない」と言われるように、男女は時として激しく争い、互いに共感することが難しい存在でもあります。人類歴史を振り返ってみると、大半が男女不平等の歴史であり、男女共同参画社会の実現に向けて今もなお様々な取り組みがなされています。男女は互いに惹かれ合っていても、ぶつかり合う存在でもある、そのようなジレンマはなぜ生じるのでしょうか?男と女、オスとメス、という性差がどのようにして生み出されるのか、またそのメカニズムがどのように進化してきたのかを研究することでその答えが見つかると思います。
地球上の全動植物のうち、54%は昆虫によって占められており「地球は昆虫の星」といっても過言ではないと言えます。昆虫が繁栄した最大の要因は、その高い適応能力にあります。不適環境を耐える休眠、ダイナミックな機能変化を生み出す変態、多様な生殖様式など、その適応戦力は人間の想像を遙かに超えるものです。また、昆虫は長い進化の歴史をもっているため、「性」についてその原型と進化過程を探るのにも適しているといえるでしょう。
そこで私達は、昆虫における多様な性決定・性分化様式に着目した研究を主要なテーマとしています。昆虫は性ホルモンを持たない、という点で脊椎動物の性決定様式と大きく異なるばかりでなく、昆虫種間でも性決定様式は多岐に渡り、多くの種においてその詳細は未解明のままです。性決定の仕組みを明らかにすることで、昆虫、ひいては動物全般における性の進化を紐解くことを目指しています。また、性分化と生殖とは密接な関係にあることから、多様な昆虫の性決定機構を理解することで昆虫の繁殖制御技術につながり、遺伝子組換え昆虫の拡散防止や害虫防除法への応用も期待できます。一方で、昆虫の高度な適応能力を支える構造・機能的基盤には性差を示すものも多くありますが、これらは生物資源として応用可能な要素が秘められていると考えられます。私達は性決定機構の解明を通じて、こうした生物資源の実用化も目指しています。






昆虫の性決定については以下も併せてご覧ください。
⇒昆虫の性決定メカニズムの謎 -スプライシングによるスイッチとは (生物の科学 遺伝, 2016年9月号掲載)

― 研究内容
研究内容については以下の動画もご参照ください
研究紹介動画 (一般の方向け)
研究紹介動画 (配属をお考えのB4、修士向け)
(1)宿主と共生細菌のせめぎ合いが生み出す性差形成機構の多様性

終齢幼虫 
各産地の成虫
節足動物はボルバキアやスピロプラズマなどの共生細菌に感染しており、これらの共生細菌は宿主の性比をメスに偏らせます。しかも、ボルバキアは宿主の性決定遺伝子に直接作用し、性差形成機構を乗っ取ることもわかってきました。この乗っ取りに対抗するため、宿主側の性決定遺伝子には多様な変異がもたらされると想像できます。私達は、性決定遺伝子に機能的な差を示すマイマイガ (Lymantria dispar japonica) 地域集団を研究材料として、性決定遺伝子に見られる地域差が共生細菌に対する対抗措置として獲得された可能性を検証します。
(2)オスとメスのせめぎ合いが生み出す性差形成機構の多様性
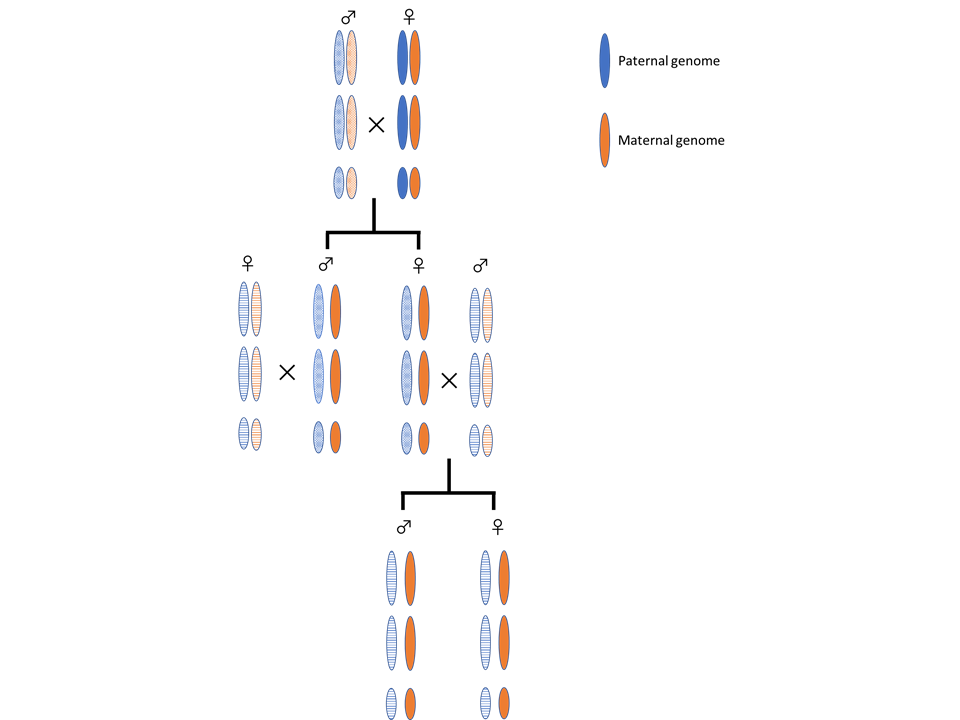
PGEによる雄親ゲノムの排除 
カツブシチャタテ
オスとメスは子孫を残す上で共同関係にありながら、いかに自分(オスであればオス、メスであればメス)の遺伝子を優先して次世代に伝えるかという点で対立関係にあります。この対立がユニークな性差形成機構を生み出します。Paternal Genome Elimination(PGE)はメスの生殖価値を高める性差形成機構のひとつです。PGEを採用する種では、母親由来の何らかの因子が父方由来の全ゲノムの不活性化を誘導し、その結果オスが産まれます。父方ゲノムは不活性化されるだけでなく、精子形成の際に捨てられてしまいます。私達はチャタテムシを用いてPGEの分子機構の解明に挑みます。PGEの解明は、ゲノムインプリンティングに関わるエピゲノム修飾やそれに伴う遺伝子発現抑制機構の理解を深めます。
(3)性差形成機構におけるミッシング・リンクの探索
団居 (まどい) 
メス成虫
哺乳類と昆虫の性差形成機構にほとんど共通点は見られません。鋏角類(クモ類)は系統発生的に哺乳類と昆虫の間に位置します。クモ類の発生学的特徴はどちらかというと脊椎動物に類似することもわかってきました。クモ類の性差形成機構の解明は、哺乳類と昆虫の性差形成機構を結ぶミッシング・リンクの発見につながる可能性があります。この点に着目し、私達はオオヒメグモ (Parasteatoda tepidariorum) を用いてクモ類の性差形成機構の解明を目指します。本研究はJT生命誌研究館 細胞・発生・進化研究室との共同研究として行っております。
(4)個体の性差はどこまでが細胞自律的に形成されるのか

雌雄モザイク (白=♂、黒=♀) 
単離細胞 
scRNA-seqデータ解析
哺乳類の性差は、血液を巡る性ホルモンの刺激によって形成されると言われていました。ところが最近の研究により、個々の細胞が自身のもつ性染色体に依存して自律的に性分化を遂げることもわかってきました。このような細胞自律的な性差形成機構は昆虫において広く認められます。では、個体の性差はどこまでが細胞自律的に形成されるのでしょうか?私達の研究分野ではオスとメスの細胞の混成体である雌雄モザイクカイコから細胞を分取し、1細胞シーケンス解析 (scRNA-seq) を行うことでこの謎を明らかにします。
本研究は科学研究費助成事業 基盤研究(B) 「昆虫の性分化はどこまで細胞自律的か?雌雄モザイク体1細胞シーケンス法による検証」のテーマとして採用されております。また、本研究の一部は東京農工大学蚕学研究室との共同研究として行っております。
(5) ポリアレリック性決定遺伝子がもたらす性スペクトラム
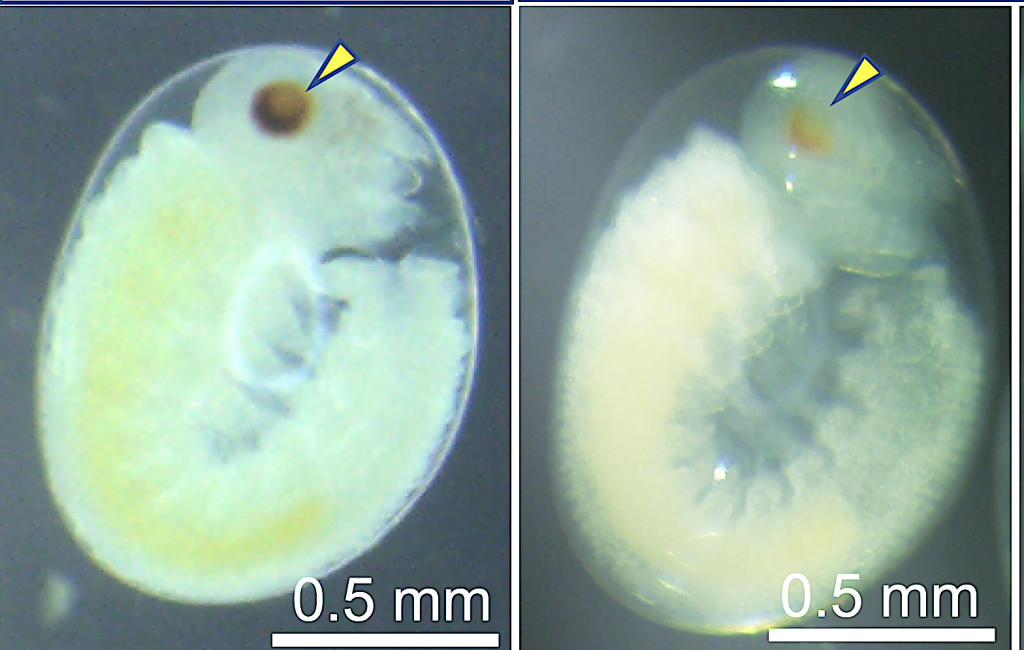
胚子 
成虫
ハチ目や一部のカメムシ目、アザミウマでは受精卵から産まれる2倍体がメス、未授精卵から産まれる半数体がオスとなる半数倍数性という性決定機構が用いられています。ハチ目では性決定遺伝子であるCsd が異なるアレルの組み合わせとなる時にのみメス化を促進することがわかっていますが、Csdがどのようにメス化を促進するかについてはほとんどわかっていません。私達はハチ目のモデル生物であるカブラハバチを用いてこの種のCsdを同定し、アレルの違いによってメス化が促進される仕組みの解明を目指します。
本研究は新学術領域研究「性スペクトラム:連続する表現型としての雌雄」(領域代表: 立花誠)の研究計画班テーマとして採用されております。また、本研究の一部は農研機構昆虫デザイン技術グループとの共同研究として行っております。